Блок 6. Эволюция живой природы
6.4. Макроэволюция. Направления и пути эволюции (А. Н. Северцов, И. И. Шмальгаузен). Биологический прогресс и регресс, ароморфоз, идиоадаптация, дегенерация. Причины биологического прогресса и регресса. Гипотезы возникновения жизни на Земле. Основные ароморфозы в эволюции растений и животных. Усложнение живых организмов на Земле в процессе эволюции.
Макроэволюция
Образование вида ознаменовывает собой новый виток эволюционного процесса, поскольку особи этого вида, будучи более приспособленными к условиям среды, нежели особи материнского вида, постепенно расселяются на новые территории, и уже в его популяциях играют свою созидательную роль мутагенез, популяционные волны, изоляция и естественный отбор. Со временем эти популяции дают начало новым видам, которые вследствие генетической изоляции имеют гораздо больше признаков сходства между собой, чем с видами того рода, от которого отпочковался вид - родоначальник, и, таким образом, возникает новый род, затем — новое семейство, отряд (порядок), класс и т. д. Совокупность эволюционных процессов, которые приводят к возникновению надвидовых таксонов (родов, семейств, отрядов, классов и т. п.), называется макроэволюцией. Макроэволюционные процессы как бы обобщают микроэволюционные изменения, происходящие в течение длительного времени, выявляя при этом основные тенденции, направления и закономерности эволюции органического мира, которые не поддаются наблюдению на более низком уровне. До сих пор никаких специфических механизмов макроэволюции не выявлено, поэтому считается, что она осуществляется только посредством микроэволюционных процессов, однако эта позиция постоянно подвергается вполне обоснованной критике.
Возникновение сложной иерархической системы органического мира в значительной степени является результатом неодинаковой скорости эволюционирования различных групп организмов. Так, уже упоминавшийся гинкго двулопастный как бы «законсервировался» на тысячи лет, тогда как достаточно близкие к нему сосны существенно изменились за это время.
Направления и пути эволюции (А. Н. Северцов, И. И. Шмальгаузен). Биологический прогресс и регресс, ароморфоз, идиоадаптация, дегенерация.
Анализируя историю органического мира, можно заметить, что в определенные промежутки времени господствовали отдельные группы организмов, которые затем клонились к упадку либо исчезали вовсе. Таким образом, можно различить три магистральных направления эволюции: биологический прогресс, биологический регресс и биологическая стабилизация. Значительный вклад в разработку учения о направлениях и путях эволюции внесли русские эволюционисты А. Н. Северцов и И. И. Шмальгаузен. Биологический прогресс связан с биологическим процветанием группы в целом и характеризует ее эволюционный успех. Он отражает закономерное развитие живой природы от простого к сложному, от более низкой степени организации к более высокой. По А. Н. Северцову, критериями биологического прогресса являются увеличение численности особей данной группы, расширение ее ареала, а также появление и развитие в ее составе групп низшего ранга (преобразование вида в род, рода в семейство и т. д.). В настоящее время биологический прогресс наблюдается у покрытосеменных растений, насекомых, костистых рыб и млекопитающих.
По А. Н. Северцову, биологический прогресс может достигаться вследствие определенных морфофизиологических преобразований организмов, при этом он выделил три основные пути достижения: арогенез, аллогенез и катагенез.
Арогенез, или морфофизиологический прогресс, связан со значительным расширением ареала данной группы организмов вследствие приобретения больших изменений строения — ароморфозов. Ароморфозом называют эволюционное преобразование строения и функций организма, которое повышает его уровень организации и открывает новые возможности для приспособления к разнообразным условиям существования.
Примерами ароморфозов являются возникновение эукариотической клетки, многоклеточности, появление сердца у рыб и разделение его полной перегородкой у птиц и млекопитающих, формирование цветка у покрытосеменных и т. п.
Аллогенез, в отличие от арогенеза, не сопровождается расширением ареала, однако внутри старого возникает значительное разнообразие форм, имеющих частные приспособления к среде обитания — идиоадаптации. Идиоадаптация — это мелкое морфофизиологическое приспособление к специальным условиям среды, полезное в борьбе за существование, но не изменяющее уровня организации. Эти изменения иллюстрируют покровительственная окраска у животных, разнообразие ротовых аппаратов у насекомых, колючки растений и др. Не менее удачным примером являются дарвиновы вьюрки, специализирующиеся на различных видах пищи, у которых преобразования вначале затронули клюв, а затем и другие части тела — оперение, хвост и т. п.
Как это ни парадоксально, но и упрощение организации может вести к биологическому прогрессу. Этот путь называется катагенезом.
Дегенерация — это упрощение организмов в процессе эволюции, которое сопровождается потерей определенных функций или органов.
Большей частью она характерна для паразитических организмов и животных, перешедших к при-крепленному способу жизни. Так, растение-паразит повилика европейская лишена хлорофилла, а ее листья редуцированы до чешуек, питается же она за счет поглощения питательных веществ из тела растения-хозяина с помощью специальных присосок. Паразитирующие в тонком кишечнике человека и других животных цепни и лентецы в процессе эволюции принесли свою пищеварительную систему и почти все органы чувств в жертву гипертрофированной половой системе, обеспечивающей необычайную плодовитость (до 11 млрд. яиц в течение жизни). Сидячий образ жизни привел у взрослых представителей одной из групп хордовых — асцидий — к исчезновению не только органов чувств и пищеварительной системы, но и самой хорды, обнаруживающейся только у личинок.
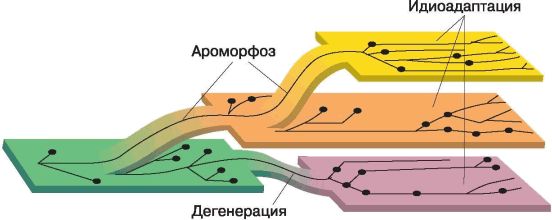
Фаза биологического прогресса сменяется фазой биологической стабилизации, сущность которой заключается в сохранении признаков данного вида как наиболее благоприятных в данном микроокружении. По И. И. Шмальгаузену, она вовсе «не означает прекращения эволюции, наоборот, означает максимальную согласованность организма с изменениями среды». В фазе биологической стабилизации находятся «живые ископаемые» латимерия, гингко и др.
Антиподом биологического прогресса является биологический регресс — эволюционный упадок данной группы вследствие невозможности приспособиться к изменениям окружающей среды. Он проявляется в снижении численности популяций, сужении ареалов, уменьшении количества групп низшего ранга в составе высшего таксона. Группе организмов, которая пребывает в состоянии биологического регресса, угрожает вымирание. В истории органического мира можно увидеть много примеров такого явления, и в настоящее время регресс характерен для некоторых папоротников, амфибий и рептилий. С появлением человека биологический регресс зачастую обусловлен его хозяйственной деятельностью.
Направления и пути эволюции органического мира не являются взаимоисключающими, то есть по-явление ароморфоза не означает, что идиоадаптация или дегенерация произойти уже не может. Напротив, согласно разработанному А. Н. Северцовым и И. И. Шмальгаузеном правилу смены фаз, различные направления эволюционного процесса и пути достижения биологического прогресса закономерно сменяют друг друга. В ходе эволюции эти пути сочетаются: достаточно редкие ароморфозы переводят группу организмов на качественно новый уровень организации, а в дальнейшем историческое развитие идет по пути идиоадаптации либо дегенерации, обеспечивающих приспособление к конкретным условиям среды обитания.
Причины биологического прогресса и регресса
В процессе эволюции планку естественного отбора преодолевают и, соответственно, прогрессируют только те группы организмов, у которых наследственная изменчивость создает достаточное количество комбинаций, которые могут обеспечить выживание группы в целом.
Те же группы, у которых по каким-то причинам не имеется такого резерва, в большинстве случаев обречены на вымирание. Зачастую это связано с низким давлением отбора на предыдущих этапах эволюционного процесса, приведшим к узкой специализации группы или даже дегенеративным явлениям. Следствием этого является невозможность приспособиться к новым условиям среды при ее резких изменениях. Ярким примером тому является внезапная гибель динозавров вследствие падения гигантского небесного тела на Землю 65 млн лет назад, которое повлекло за собой землетрясение, поднятие в воздух миллионов тонн пыли, резкое похолодание, гибель боль¬шей части растений и растительноядных животных. В то же время предки современных млекопитающих, не имея узких предпочтений источников питания и будучи теплокровными, смогли пережить эти условия и занять на планете господствующее положение.
Гипотезы возникновения жизни на Земле
Из всего спектра гипотез образования Земли наибольшее количество фактов свидетельствует в пользу теории «Большого взрыва». Ввиду того, что данное научное допущение зиждется в основном на теоретических расчетах, подтвердить его экспериментально призван большой адронный коллайдер, сооруженный в Европейском центре ядерных исследований вблизи г. Женева (Швейцария). Согласно теории «Большого взрыва», Земля образовалась свыше 4,5 млрд лет назад вместе с Солнцем и другими планетами Солнечной системы в результате конденсации газопылевого облака. Снижение температуры планеты и миграция химических элементов на ней способствовали ее расслоению на ядро, мантию и кору, а происходившие затем геологические процессы (движение тектонических плит, вулканическая деятельность и т. д.) стали причиной формирования атмосферы и гидросферы.
Жизнь существует на Земле также очень давно, о чем свидетельствуют ископаемые остатки разнообразных организмов в горных породах, однако физические теории не могут дать ответа на вопрос о времени и причинах ее возникновения. Существуют две противоположные точки зрения на возникновение жизни на Земле: теории абиогенеза и биогенеза. Теории абиогенеза утверждают возможность происхождения живого из неживого. К ним относят креационизм, гипотезу самозарождения и теорию биохимической эволюции А. И. Опарина.
Фундаментальным положением креационизма являлось сотворение мира неким сверхъестественным существом (Творцом), что нашло свое отражение в мифах народов мира и религиозных культах, однако возраст планеты и жизни на ней намного превышает указанные в этих источниках сроки, да и несоответствий в них предостаточно.
Основателем теории самозарождения жизни считается древнегреческий ученый Аристотель, который утверждал, что возможно многократное появление новых существ, например, дождевых червей из луж, а червей и мух — из гнилого мяса. Однако эти воззрения были опровергнуты в XVII-XIX веках смелыми опытами Ф. Реди и Л. Пастера.
Итальянский врач Франческо Реди в 1688 году поместил кусочки мяса в горшки и плотно запечатал их, однако никаких червей в них не завелось, тогда как в открытых горшках они по¬явились. Дабы опровергнуть бытовавшее тогда убеждение, что жизненное начало содержится в воздухе, он повторил свои опыты, однако горшки не запечатал, а закрыл несколькими слоями кисеи, и вновь жизнь не появилась. Несмотря на убедительные данные, полученные Ф. Реди, исследования А. ван Левенгука дали новую пищу для дискуссий о «жизненном начале», которые продолжились в течение всего следующего века.
Другой итальянский исследователь — Ладзаро Спалланцани — в 1765 году видоизменил опыты Ф. Реди, прокипятив в течение нескольких часов мясные и овощные отвары и запечатав их. По прошествии нескольких дней он также не обнаружил там никаких признаков жизни и сделал вывод, что живое может возникнуть только от живого.
Последний удар теории спонтанного самозарождения нанес великий французский микробиолог Луи Пастер в 1860 году, поместивший прокипяченный бульон в колбу с S-образным горлышком и не получивший никаких зародышей. Казалось бы, это свидетельствовало в пользу теорий биогенеза, однако оставался открытым вопрос о том, каким же путем возник самый-самый первый организм.
Ответить на него попытался советский биохимик А. И. Опарин, пришедший к выводу о том, что состав атмосферы Земли на первых этапах ее существования был совсем не таким, как в наше время. Скорее всего, она состояла из аммиака, метана, углекислого газа и водяных паров, но не содержала свободного кислорода. Под действием электрических разрядов высокой мощности и при высокой температуре в ней могли синтезироваться простейшие органические соединения, что и было подтверждено экспериментами С. Миллера и Г. Юри в 1953 году, получивших из вышеупомянутых соединений несколько аминокислот, простые углеводы, аденин, мочевину, а также простейшие жирные, муравьиную и уксусную кислоты. Тем не менее синтез органических веществ еще не означает возникновения жизни, поэтому А. И. Опарин выдвинул гипотезу биохимической эволюции, согласно которой разнообразные органические вещества возникали и объединялись в более крупные молекулы на мелководьях морей и океанов, где условия для химического синтеза и полимеризации являются наиболее благоприятными.
Некоторые из этих веществ постепенно образовывали в воде устойчивые комплексы — коацерваты, или коацеватные капли,напоминающие капли жира в бульоне. В эти коацерваты по¬ступали разнообразные вещества из окружающего раствора, которые подвергались химическим превращениям, происходящим в каплях. Как и органические вещества, коацерваты сами по себе не являлись живыми существами, а были очередной ступенью в их возникновении.
Те из коацерватов, которые имели удачное соотношение веществ в своем составе, в особенности белки и нуклеиновые кислоты, благодаря каталитическим свойствам белков-ферментов со временем приобрели способность воспроизводить себе подобных и осуществлять реакции обмена веществ, при этом структуру белков кодировали нуклеиновые кислоты.
Однако, помимо размножения, для живых систем характерна зависимость от поступления энергии извне. Эта проблема первоначально решалась за счет бескислородного расщепления органических веществ из окружающей среды (кислорода в атмосфере на тот момент не было), т. е. гетеротрофного питания. Некоторые из поглощаемых органических веществ оказались способными аккумулировать энергию солнечного света, как, например, хлорофилл, что дало возможность ряду организмов перейти к автотрофному питанию. Выделение кислорода в атмосферу в процессе фотосинтеза привело к появлению более эффективного кислородного дыхания, возникновению озонового слоя и, в конечном итоге, выходу организмов на сушу. Таким образом, результатом химической эволюции явилось появление протобионтов — первичных живых организмов, от которых в результате биологической эволюции произошли все существующие в настоящее время виды.
Теория биохимической эволюции в наше время является наиболее подтвержденной, однако представление о конкретных механизмах возникновения жизни изменились. Например, выяснилось, что образование органических веществ начинается еще в космосе, а органические вещества играют важную роль даже в самом образовании планет, обеспечивая слипание мелких частей. Также формирование органических веществ происходит и в недрах планеты: при одном извержении вулкан выбрасывает до 15 т органики. Существуют и другие гипотезы относительно механизмов концентрирования органических веществ: замораживания раствора, абсорбции (связывания) на поверхности определенных минеральных соединений, действия природных катализаторов и т. п.
Возникновение жизни на Земле в настоящее время невозможно, поскольку любые органические вещества, спонтанно образовавшиеся в любой точке планеты, тотчас же были бы окислены свободным кислородом атмосферы или использованы гетротрофными организмами. Это понимал еще в 1871 году Ч. Дарвин. Теории биогенеза отрицают самопроизвольное зарождение жизни. Основными из них являются гипотеза стационарного состояния и гипотеза панспермии. Первая из них базируется на том, что жизнь существует вечно, тем не менее, на нашей планете есть очень древние породы, в которых следы деятельности органического мира отсутствуют.
Гипотеза панспермии утверждает, что зародыши жизни были занесены на Землю из космоса некими пришельцами либо божественным провидением. В пользу этой гипотезы свидетельствуют два факта: необходимость для всего живого достаточно редкого на планете, но часто встречающегося в метеоритах молибдена, а также находка организмов, похожих на бактерии, на метеоритах с Марса. Однако каким образом жизнь возникла на других планетах, остается невыясненным.
Основные ароморфозы в эволюции растений и животных
Растительные и животные организмы, представляющие различные ветви эволюции органического мира, в процессе исторического развития независимо приобретали определенные черты строения, которые будут охарактеризованы далее.
У растений важнейшими из них являются переход от гаплоидности к диплоидности, независимость от воды в процессе оплодотворения, переход от наружного оплодотворения к внутреннему и возникновение двойного оплодотворения, расчленение тела на органы, развитие проводящей си-стемы, усложнение и усовершенствование тканей, а также специализация опыления с помощью насекомых и распространения семян и плодов.
Переход от гаплоидности к диплоидности сделал растения более устойчивыми к действию факторов окружающей среды вследствие снижения риска проявления рецессивных мутаций. По- видимому, это преобразование коснулось предков сосудистых растений, в число которых не входят моховидные, характеризующиеся преобладанием в жизненном цикле гаметофита.
Главные ароморфозы в эволюции животных связаны с возникновением многоклеточности и все большим расчленением всех систем органов, возникновением прочного скелета, развитием центральной нервной системы, а также общественного поведения в различных группах высоко-организованных животных, что дало толчок и к прогрессу человека.
Усложнение живых организмов в процессе эволюции
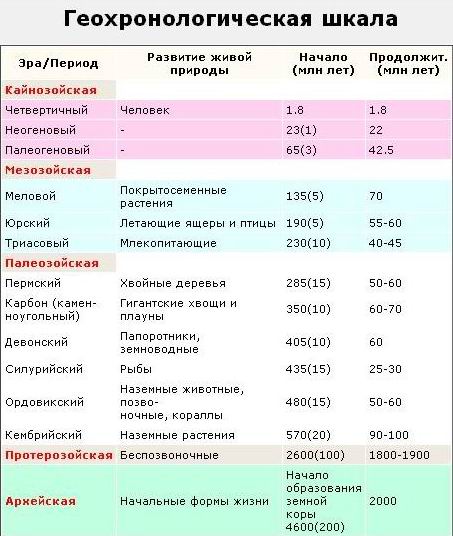
Историю органического мира на Земле изучают по сохранившимся остаткам, отпечаткам и другим следам жизнедеятельности живых организмов. Она является предметом науки палеонтологии. Исходя из того, что остатки разных организмов расположены в различных пластах горных пород, была создана геохронологическая шкала, согласно которой историю Земли разделили на определенные промежутки времени: зоны, эры, периоды и века.
Эоном называют большой промежуток времени в геологической истории, объединяющий несколько эр. В настоящее время выделяют только два зона: криптозой (скрытая жизнь) и фанерозой (явная жизнь). Эра — это промежуток времени в геологической истории, являющийся подразделением эона, объединяющий, в свою очередь, периоды. В криптозое выделяют две эры (архей и протерозой), тогда как в фанерозое — три (палеозой, мезозой и кайнозой).
Важную роль в создании геохронологической шкалы сыграли руководящие ископаемые — остатки организмов, которые были многочисленны в определенные промежутки времени и хорошо сохранились. Развитие жизни в криптозое. Архей и протерозой составляют большую часть истории жизни (период 4,6 млрд. лет — 0,6 млрд. лет назад), однако сведений о жизни в тот период недостаточно. Первые остатки органических веществ биогенного происхождения имеют возраст около 3,8 млрд. лет, а прокариотические организмы существовали уже 3,5 млрд. лет назад. Первые прокариоты входили в состав специфических экосистем — цианобактериальных матов, благо¬даря деятельности которых образовались специфические осадочные породы строматолиты («каменные ковры»).
Понять жизнь давних прокариотических экосистем помогло открытие их современных аналогов — строматолитов в заливе Шарк-Бей в Австралии и специфических пленок на поверхности почвы в заливе Сиваш в Украине. На поверхности цианобактериальных матов расположены фото- синтезирующие цианобактерии, а под их слоем — чрезвычайно разнообразные бактерии других групп и архей. Минеральные вещества, которые оседают на поверхность мата и образовываются за счет его жизнедеятельности, откладываются пластами (приблизительно 0,3 мм в год). Такие примитивные экосистемы могут существовать лишь в непригодных для жизни других организмов местах, и действительно, оба вышеупомянутые местообитания характеризуются чрезвычайно высокой соленостью.
Многочисленные данные свидетельствуют о том, что поначалу Земля имела атмосферу возобновляемого характера, в состав которой входили: углекислый газ, водяной пар, оксид серы, а также угарный газ, водород, сероводород, аммиак, метан и т. п. Первые организмы Земли были анаэробами, однако благодаря фотосинтезу цианобактерий в среду выделялся свободный кислород, который сначала быстро связывался с восстановителями, находящимися в среде, и лишь после связывания всех восстановителей среда начала приобретать окислительные свойства. О таком переходе свидетельствуют отложение окисленных форм железа — гематита и магнетита.
Около 2 млрд. лет назад в результате геофизических процессов практически все несвязанное в осадочных породах железо переместилось к ядру планеты, а кислород начал накапливаться в атмосфере из-за отсутствия этого элемента — произошла «кислородная революция». Она явилась переломным этапом в истории Земли, который повлек за собой не только смену состава атмосферы и образование озонового экрана в атмосфере — главной предпосылки для заселения суши, но и состава пород, формирующихся на поверхности Земли. В протерозое произошло и другое важное событие — возникновение эукариот. В последние годы удалось собрать убедительные доказательства теории эндосимбиогенетического происхождения эукариотической клетки — путем симбиоза нескольких прокариотических клеток. Вероятно, «главным» предком эукариот стали архей, которые перешли к поглощению пищевых частиц путем фагоцитоза. Наследственный аппарат переместился вглубь клетки, сохранив, тем не менее, связь с мембраной благодаря переходу внешней мембраны возникшей ядерной оболочки в мембраны эндоплазматической сети.
Поглощенные клеткой бактерии могли не перевариваться, а оставаться живыми и продолжать свое функционирование. Считают, что митохондрии ведут свое происхождение от пурпурных бактерий, утративших способность к фотосинтезу и перешедших к окислению органических веществ. Симбиоз с другими фотосинтезирующими клетками привел к возникновению пластид у растительных клеток. Вероятно, жгутики эукариотических клеток возникли вследствие симбиоза с бактериями, которые, подобно современным спирохетам, были способны к извивающимся движениям. Поначалу наследственный аппарат эукариотических клеток был устроен приблизи-тельно так же, как у прокариот, и лишь позднее, вследствие необходимости управления большой и сложной клеткой, образовались хромосомы. Геномы внутриклеточных симбионтов (митохондрий, пластид и жгутиков) в целом сохранили прокариотическую организацию, но большая часть их функций перешла к ядерному геному. Эукариотические клетки возникали неоднократно и независимо друг от друга. Например, красные водоросли возникли в результате симбиогенеза с цианобактериями, а зеленые водоросли — с бактериями-прохлорофитами.
Остальные одномембранные органеллы и ядро эукариотической клетки, согласно эндомембранной теории, возникли из впячиваний мембраны прокариотической клетки.
Точное время появления эукариот неизвестно, поскольку уже в отложениях возрастом около 3 млрд лет присутствуют отпечатки клеток, имеющих похожие размеры. Точно эукариоты за¬фиксированы в породах возрастом около 1,5-2 млрд лет, но только после кислородной революции (около 1 млрд лет назад) сложились условия, благоприятные для них.
В конце протерозойской эры (не менее 1,5 млрд лет назад) уже существовали и многоклеточные эукариотические организмы. Многоклеточность, как и эукариотическая клетка, неоднократно возникала у разных групп организмов.
Существуют различные взгляды на происхождение многоклеточных животных. По одним данным их родоначальниками были многоядерные, подобные инфузориям, клетки, которые затем распались на отдельные одноядерные клетки.
Другие гипотезы связывают происхождение многоклеточных животных с дифференцировкой клеток колониальных одноклеточных. Расхождения между ними касаются возникновения слоев клеток у первоначального многоклеточного животного. Согласно гипотезы гастреи Э. Геккеля, то происходит путем впячивания одной из стенок однослойного многоклеточного организма, как у кишечнополостных. В противовес ей И. И. Мечников сформулировал гипотезу фагоцителлы, считая предками многоклеточных однослойные шарообразные колонии наподобие вольвокса, которые поглощали пищевые частицы путем фагоцитоза. Клетка, захватившая частицу, теряла жгутик и переходила вглубь организма, где и осуществляла пищеварение, а по окончании процесса возвращалась на поверхность. Со временем произошло разделение клеток на два слоя с определенными функциями — внешний обеспечивал движение, а внутренний — фагоцитоз. Такой организм И. И. Мечников назвал фагоцителлой.
В течение продолжительного времени многоклеточные эукариоты проигрывали в конкурентной борьбе прокариотическим организмам, однако в конце протерозоя (800-600 млн лет тому) вследствие резкого изменения условий на Земле — снижения уровня морей, роста концентрации кислорода, уменьшения концентрации карбонатов в морской воде, регулярных циклов похолодания — многоклеточные эукариоты получили преимущества над прокариотами. Если до этого времени встречались только отдельные многоклеточные растения и, возможно, грибы, то с этого момента в истории Земли известны и животные. Из возникших в конце протерозоя фаун лучше других изучены эдиакарская и вендская. Животных вендского периода принято включать в состав особой группы организмов или относить к таким типам, как кишечнополостные, плоские черви, членистоногие и др. Однако ни у одной из этих групп нет скелетов, что может свидетельствовать об отсутствии хищников.
Развитие жизни в палеозойской эре. Палеозойская эра, длившаяся более 300 млн лет, делится на шесть периодов: кембрийский, ордовикский, силурийский, девонский, каменноугольный (карбон) и пермский. В кембрийском периоде суша состояла из нескольких материков, расположенных преимущественно в Южном полушарии. Самыми многочисленными фотосинтезирующими организмами в этот период были цианобактерии и красные водоросли. В толще воды жили фораминиферы и радиолярии. В кембрии появляется огромное количество скелетных животных организмов, о чем свидетельствуют многочисленные ископаемые остатки. Эти организмы относились примерно к 100 типам многоклеточных животных, как современным (губки, кишечнополостные, черви, членистоногие, моллюски), так и исчезнувшим, например: огромный хищник аномалокарис и колониальные граптолиты, которые плавали в толще воды или были прикреплены ко дну. Суша на протяжении кембрия оставалась почти незаселенной, однако процесс почвообразования уже начали бактерии, грибы и, возможно, лишайники, а в конце периода на сушу вышли малощетинковые черви и многоножки.
В ордовикском периоде уровень вод Мирового океана поднялся, что привело к затоплению материковых низменностей. Основными продуцентами в этот период были зеленые, бурые и красные водоросли. В отличие от кембрия, в котором рифы строили губки, в ордовике их сменяют коралловые полипы. Расцвет переживали брюхоногие и головоногие моллюски, а также трилобиты (ныне вымершие родственники паукообразных). В этом периоде впервые зафиксированы и хордовые, в частности бесчелюстные. В конце ордовика произошло грандиозное вымирание, которое уничтожило около 35 % семейств и более 50 % родов морских животных.
Силурийский период характеризуется усилением горообразования, которое привело к осушению материковых платформ. Ведущую роль в фауне беспозвоночных силура играли головоногие моллюски, иглокожие и гигантские ракоскорпионы, тогда как среди позвоночных сохраняется большое разнообразие бесчелюстных и появляются рыбы. В конце периода на сушу вышли первые сосудистые растения — риниофиты и плауновидные, которые начали колонизацию мелководья и приливно-отливной зоны побережий. На сушу вышли и первые представители класса паукообразные.
В девонском периоде вследствие поднятия суши образовались большие мелководья, которые пересыхали и даже промерзали, поскольку климат становился еще более континентальным, чем в силуре. В морях преобладают кораллы и иглокожие, тогда как головоногие моллюски представ¬лены спирально закрученными аммонитами. Среди позвоночных девона расцвета достигли рыбы, причем на смену панцирным пришли и хрящевые, и костные, а также двоякодышащие и кистеперые. В конце периода появляются первые амфибии, которые сначала жили в воде.
В среднем девоне на суше появились первые леса из папоротников, плаунов и хвощей, которые были заселены червями и многочисленными членистоногими (многоножками, пауками, скорпионами, бескрылыми насекомыми). В конце девона появились первые голосеменные. Освоение суши растениями привело к уменьшению выветривания и усилению почвообразования. Закрепление почв привело к возникновению русел рек.
В каменноугольном периоде суша была представлена двумя материками, разделенными океаном, а климат стал заметно более теплым и влажным. К концу периода произошло небольшое поднятие суши, а климат сменился более континентальным. В морях господствовали фораминиферы, кораллы, иглокожие, хрящевые и костные рыбы, а пресные водоемы населяли двухстворчатые моллюски, ракообразные и разнообразные земноводные. В середине карбона возникли мелкие насекомоядные рептилии, а среди насекомых появились крылатые (тараканы, стрекозы).
Для тропиков были характерны заболоченные леса, в которых доминировали гигантские хвощи, плауны и папоротники, отмершие остатки которых образовали впоследствии залежи каменного угля. В середине периода в умеренной зоне, благодаря их независимости от воды в процессе оплодотворения и наличию семени, началось распространение голосеменных.
Пермский период отличался слиянием всех материков в единый суперконтинент Пангею, отступлением морей и усилением континентальности климата до такой степени, что во внутренних районах Пангеи образовались пустыни. К концу периода на суше почти исчезли древовидные папоротники, хвощи и плауны, а господствующее положение заняли засухоустойчивые голосеменные.
Несмотря на то, что крупные амфибии еще продолжали существовать, возникли разные группы рептилий, в том числе крупных растительноядных и хищных. В конце перми произошло самое большое вымирание в истории жизни, так как исчезли многие группы кораллов, трилобиты, большинство головоногих, рыб (в первую очередь хрящевых и кистеперых), а также амфибий. Морская фауна потеряла при этом 40-50% семейств и около 70% родов.
Развитие жизни в мезозое. Мезозойская эра продолжалась около 165 млн лет и характеризовалась поднятием суши, интенсивным горообразованием и снижением влажности климата. Она делится на три периода: триасовый, юрский и меловой.
В начале триасового периода климат был засушливым, однако позднее вследствие поднятия уровня морей он стал более влажным. Среди растений преобладали голосеменные, папоротники и хвощи, однако древесные формы споровых практически полностью вымерли. Высокого развития достигли некоторые кораллы, аммониты, новые группы фораминифер, двухстворчатых моллюсков и иглокожих, тогда как разнообразие хрящевых рыб уменьшилось, изменились и группы костных рыб. Господствовавшие на суше рептилии начали осваивать и водную среду, как ихтиозавры и плезиозавры. Из пресмыкающихся триаса до нашего времени дожили крокодилы, гаттерии и черепахи. В конце триаса появились динозавры, млекопитающие и птицы. В юрском периоде суперконтинент Пангея раскололся на несколько меньших. Большая часть юры была очень влажной, а к его концу климат стал более засушливым. Доминирующей группой растений были голосеменные, из которых от того времени сохранились секвойи. В морях процветали моллюски (аммониты и белемниты, двухстворчатые и брюхоногие), губки, морские ежи, хрящевые и костные рыбы. Крупные амфибии практически полностью вымерли в юрском периоде, однако появились современные группы земноводных (хвостатые и бесхвостые) и чешуйчатых (ящериц и змей), возросло разнообразие млекопитающих. К концу периода возникли и возможные предки первых птиц — археоптериксы. Однако во всех экосистемах доминировали пресмыкающиеся — ихтиозавры и плезиозавры, динозавры и летающие ящеры — птерозавры. Меловой период получил название в связи с образованием мела в осадочных породах того времени. На всей Земле, кроме приполярных областей, был стойкий теплый и влажный климат. В этом периоде возникли и приобрели широкое распространение покрытосеменные, вытеснявшие голосеменных, что повлекло за собой резкое увеличение разнообразия насекомых. В морях, по¬мимо моллюсков, костистых рыб, плезиозавров, вновь появилось огромное количество фораминифер, раковинки которых и образовали залежи мела, а на суше преобладали динозавры. Лучше приспособленные к воздушной среде птицы начали постепенно вытеснять летающих ящеров.
В конце периода произошло глобальное вымирание, в результате которого исчезли аммониты, белемниты, динозавры, птерозавры и морские ящеры, древние группы птиц, а также некоторые голосеменные. С лица Земли в целом исчезло около 16% семейств и 50% родов животных. Кризис в конце мела связывают с падением большого метеорита в Мексиканский залив, однако он, скорее всего, не был единственной причиной глобальных изменений. В ходе последующего похолодания выжили только небольшие рептилии и теплокровные млекопитающие.
Развитие жизни в кайнозое. Кайнозойская эра началась около 66 млн лет назад и продолжается до настоящего времени. Она характеризуется господством насекомых, птиц, млекопитающих и покрытосеменных растений. Кайнозой делят на три периода — палеоген, неоген и антропоген — последний из которых является самым коротким в истории Земли.
В раннем и среднем палеогене климат оставался теплым и влажным, к концу периода стало прохладнее и суше. Доминирующей группой растений стали покрытосеменные, однако, если в начале периода преобладали вечнозеленые леса, то в конце появилось много листопадных, а в засушливых зонах образовались степи. Среди рыб господствующее положение заняли костистые рыбы, а количество видов хрящевых, не-смотря на их заметную роль в соленых водоемах, незначительно. На суше из рептилий сохранились только чешуйчатые, крокодилы и черепахи, тогда как млекопитающие заняли большую часть их экологических ниш. В середине периода появились основные отряды млекопитающих, в том числе насекомоядные, хищные, ластоногие, китообразные, копытные и приматы. Изоляция материков сделала фауну и флору географически более разнообразными: Южная Америка и Австралия стали центрами развития сумчатых, а другие материки — плацентарных млекопитающих.
Неогеновый период. Земная поверхность в неогене приобрела современный вид. Климат стал более прохладным и сухим. В неогене уже сформировались все отряды современных млекопитающих, а в африканских саванах возникло семейство Гоминид и род Человек. К концу периода в приполярных областях континентов распространились хвойные леса, появились тундры, а степи умеренного пояса заняли злаки. Четвертичный период (антропоген) характеризуется периодическими сменами оледенений и потеплений. Во время оледенений высокие широты покрывались ледниками, резко снижался уровень океана, суживались тропический и субтропический пояса. На близлежащих к ледникам территориях устанавливался холодный и сухой климат, который способствовал формированию холодоустойчивых групп животных — мамонтов, гигантских оленей, пещерных львов и др. Сопутствовавшее процессу оледенения снижение уровня Мирового океана привело к образованию сухопутных мостов между Азией и Северной Америкой, Европой и Британскими островами и т. д. Миграции животных, с одной стороны, привели к взаимообогащению флор и фаун, а с другой, к вытеснению реликтов пришельцами, например, сумчатых и копытных в Южной Америке. Эти процессы, однако, не затронули Австралию, оставшуюся изолированной.
В целом, периодические изменения климата привели к формированию чрезвычайно обильного видового разнообразия, характерного для нынешнего этапа эволюции биосферы, а также оказали влияние на эволюцию человека. На протяжении антропогена несколько видов рода Человек расселились из Африки в Евразию. Около 200 тысяч лет назад в Африке возник вид Человек разумный, который после продолжительного периода существования в Африке около 70 тысяч лет назад вышел в Евразию и около 35-40 тыс. лет назад — в Америку. После периода сосуществования с близкородственными видами он вытеснил их и расселился по всей территории земного шара.
Около 10 тыс. лет назад хозяйственная деятельность человека в умеренно теплых областях земного шара начала оказывать влияние как на облик планеты (распашка земель, выжигание лесов, перевыпас пастбищ, опустынивание и т. д.), так и на животный и растительный мир вследствие сокращения ареалов их обитания и истребления, и вступил в действие антропогенный фактор. |


 Меню сайта
Меню сайта Категории раздела
Категории раздела Вход
Вход Поиск
Поиск Друзья сайта
Друзья сайта Наш опрос
Наш опрос Статистика
Статистика
