Блок 6. Эволюция живой природы
6.1. Вид, его критерии. Популяция – структурная единица вида и элементарная единица эволюции. Микроэволюция. Образование новых видов. Способы видообразования. Сохранение многообразия видов как основа устойчивости биосферы.
Вид, его критерии
Основатель современной систематики К. Линней рассматривал вид как группу сходных по морфологическим признакам организмов, которые свободно скрещиваются между собой. По мере развития биологии были получены свидетельства того, что различия между видами намного глубже, и затрагивают химический состав и концентрацию веществ в тканях, направление и скорость химических реакций, характер и интенсивность процессов жизнедеятельности, число и форму хромосом, т. е. вид является наименьшей группой организмов, отражающей их близкое родство. К тому же виды не существуют вечно — они возникают, развиваются, дают начало новым видам и исчезают.
Вид — это совокупность особей, сходных по строению и особенностям процессов жизнедеятельности, имеющих общее происхождение, свободно скрещивающихся между собой в природе и дающих плодовитое потомство.
Все особи одного вида имеют одинаковый кариотип и занимают в природе определенную географическую область — ареал.
Признаки сходства особей одного вида называют критериями вида. Так как ни один из критериев не является абсолютным, для правильного определения вида необходимо использовать совокупность критериев. Основными критериями вида являются морфологический, физиологический, биохимический, экологический, географический, этологический (поведенческий) и генетический.
1. Морфологический — совокупность внешних и внутренних признаков организмов одного вида. Несмотря на то, что у некоторых видов существуют уникальные признаки, с помощью одних только морфологических черт часто очень трудно различить близкородственные виды. Так, в последнее время открыт ряд видов-двойников, обитающих на одной территории, например домовая и курганчиковая мыши, поэтому использовать исключительно морфологический критерий для определения вида недопустимо.
2. Физиологический — сходство процессов жизнедеятельности у организмов, в первую очередь, размножения. Он также не является универсальным, поскольку некоторые виды в природе скрещиваются и дают плодовитое потомство.
3. Биохимический — сходство химического состава и протекания процессов обмена веществ. Несмотря на то, что данные показатели могут значительно варьировать у разных особей одного вида, в настоящее время им уделяется большое внимание, поскольку особенности строения и состава биополимеров помогают идентифицировать виды даже на молекулярном уровне и устанавливать степень их родства.
4. Экологический — различие видов по их принадлежности к определенным экосистемам и экологическим нишам, которые они занимают. Однако многие неродственные виды занимают сходные экологические ниши, поэтому данный критерий может быть использован для выделения вида только в совокупности с другими признаками.
5. Географический — существование популяции каждого вида в определенной части биосферы — ареале, который отличается от ареалов всех остальных видов. В связи с тем, что у множества видов границы ареалов совпадают, а также имеется ряд видов-космополитов, ареал которых охватывает огромные пространства, географический критерий также не может служить маркерным «видовым» признаком.
6. Генетический — постоянство признаков хромосомного набора — кариотипа — и нуклеотидного состава ДНК у особей одного вида. В связи с тем, что негомологичные хромосомы не могут конъюгировать во время мейоза, потомство от скрещивания особей разных видов с неодинаковым набором хромосом либо не появляется вовсе, либо не плодовито. Это создает репродуктивную изоляцию вида, поддерживает его целостность и обеспечивает реальность существования в природе. Данное правило может нарушаться в случае скрещивания близких по происхождению видов с одинаковым кариотипом или возникновения различных мутаций, однако исключение лишь подтверждает общее правило, и виды следует рассматривать как устойчивые генетические системы. Генетический критерий является основным в системе критериев вида, но также не исчерпывающим.
При всей сложности системы критериев вид нельзя представить как группу абсолютно идентичных по всем параметрам организмов, то есть клонов. Наоборот, для многих видов характер¬но значительное разнообразие даже внешних признаков, как, например, для одних популяций божьих коровок характерно преобладание в окраске красного цвета, а для других — черного.
Популяция — структурная единица вида и элементарная единица эволюции.
Трудно представить, чтобы в реальности особи одного вида были равномерно распределены по земной поверхности в пределах ареала, поскольку, например, лягушка озерная обитает в основ¬ном в достаточно редких стоячих пресных водоемах, и вряд ли ее можно встретить на полях и в лесах. Виды в природе чаще всего распадаются на отдельные группы, в зависимости от подходящих по совокупности условий местам обитания — популяции.
Популяция — группа особей одного вида, занимающих часть его ареала, свободно скрещивающихся между собой и относительно обособленных от других совокупностей особей того же вида в течение более или менее длительного времени.
Популяции могут быть разделены не только пространственно, они могут обитать даже на одной территории, но иметь различия в пищевых предпочтениях, сроках размножения и т. д.
Таким образом, вид — это совокупность популяций особей, обладающих рядом общих морфологических, физиологических, биохимических признаков и типов взаимоотношений с окружающей средой, населяющих определенный ареал, а также способных скрещиваться между собой с образованием плодовитого потомства, но почти или совсем не скрещивающихся с другими группами особей того же вида.
Внутри видов с большими ареалами, охватывающими территории с разными условиями жизни, иногда различают и подвиды — большие популяции или группы соседних популяций, имеющих стойкие морфологические отличия от других популяций.
Популяции разбросаны по земной поверхности не случайным образом, они привязаны к конкретным ее участкам. Совокупность всех факторов неживой природы, необходимых для проживания особей данного вида, называется местообитанием. Однако только этих факторов может быть недостаточно для занятия этого участка популяцией, поскольку она должна быть еще вовлечена в тесное взаимодействие с популяциями других видов, то есть занять определенное место в сообществе живых организмов — экологическую нишу. Так, австралийский сумчатый медведь коала при всех прочих равных условиях не может существовать без своего основного источника питания — эвкалипта.
Образующие неразрывное единство в одних и тех же местообитаниях популяции различных видов обыкновенно обеспечивают более или менее замкнутый круговорот веществ и являются элементарными экологическими системами (экосистемами) — биогеоценозами.
При всей своей требовательности к условиям окружающей среды популяции одного вида не-однородны по занимаемой площади, численности, плотности и пространственному размещению особей, часто образующих более мелкие группы (семьи, стаи, стада и др.), полу, возрасту, генофонду и т. д., поэтому различают их размерную, возрастную, половую, пространственную, генетическую, этологическую и другие структуры, а также динамику.
Важными характеристиками популяции являются генофонд — совокупность генов, характерных для особей данной популяции или вида, а также частоты определенных аллелей и генотипов. Разные популяции одного и того же вида изначально имеют неодинаковый генофонд, так как новые территории осваивают особи со случайными, а не специально подобранными генами. Под действием внутренних и внешних факторов генофонд подвергается еще более существенным изменениям: обогащается за счет возникновения мутаций и нового сочетания признаков и обедняется в результате выпадения отдельных аллелей при гибели или миграции некоторого числа особей.
Новые признаки и их сочетания могут быть полезными, нейтральными или вредными, поэтому в популяции выживают и успешно размножаются только приспособленные к данным условиям окружающей среды особи. Однако в двух различных точках земной поверхности условия окружающей среды никогда не бывают полностью идентичными, поэтому и направление изменений даже в двух соседних популяциях может быть совершенно противоположным или они будут протекать с различной скоростью. Результатом изменений генофонда является расхождение популяций по морфологическим, физиологическим, биохимическим и другим признакам. Если популяции при этом еще и изолированы друг от друга, то они могут давать начало новым видам.
Так, возникновение каких-либо препятствий в скрещивании особей различных популяций одного вида, например, вследствие образования горных массивов, изменения русел рек, различий в сроках размножения и т. д., приводит к тому, что популяции постепенно приобретают все больше различий и, в конце концов, становятся различными видами. Некоторое время на границах этих популяций происходит скрещивание особей и возникают гибриды, однако со временем и эти контакты исчезают, т. е. популяции из открытых генетических систем становятся закрытыми.
Несмотря на то, что действию факторов окружающей среды подвергаются в первую очередь отдельные особи, изменение генетического состава у единичного организма является незначительным и проявится в лучшем случае только у его потомков. Подвиды, виды и более крупные таксоны также не подходят на роль элементарных единиц эволюции, поскольку они не отличаются морфологическим, физиологическим, биохимическим, экологическим, географическим и генетическим единством, тогда как популяции как наименьшие структурные единицы вида, накапливающие разнообразие случайных изменений, худшие из которых будут отсеяны, соответствуют этому условию и являются элементарными единицами эволюции.
Микроэволюция
Изменение генетической структуры популяций не всегда приводит к образованию нового вида, а может лишь улучшать приспособление популяции к конкретным условиям среды, однако виды не являются вечными и неизменными — они способны развиваться. Этот процесс необратимого исторического изменения живого называется эволюцией. Первично эволюционные преобразования происходят внутри вида на уровне популяций. В их основе лежат, прежде всего, мутационный процесс и естественный отбор, приводящие к изменению генофонда популяций и вида в целом, или даже к образованию новых видов. Совокупность этих элементарных эволюционных событий называется микроэволюцией.
Популяции характеризуются огромным генетическим разнообразием, которое часто не про¬является фенотипически. Генетическое разнообразие возникает вследствие спонтанного мутагенеза, который происходит непрерывно. Большинство мутаций неблагоприятны для организма и снижают жизнеспособность популяции в целом, но, если они рецессивны, то могут сохраняться в гетерозиготе в течение продолжительного времени. Некоторые мутации, не имеющие приспособительной ценности в данных условиях существования, способны приобрести такую ценность в будущем или при освоении новых экологических ниш, создавая, таким образом, резерв наследственной изменчивости.
Значительное влияние на микроэволюционные процессы оказывают колебания численности особей в популяциях, миграции и катастрофы, а также изоляция популяций и видов.
Новый вид является промежуточным результатом эволюции, но никак не ее итогом, поскольку на этом микроэволюция не прерывается — она продолжается далее. Возникающие новые виды в случае удачного сочетания признаков заселяют новые местообитания, и, в свою очередь, дают начало новым видам. Такие группы близкородственных видов объединяются в роды, семейства и т. д. Эволюционные процессы, происходящие в надвидовых группах, называются уже макроэволюцией. В отличие от макроэволюции, микроэволюция протекает в гораздо более сжатые сроки, тогда как первой требуются десятки и сотни тысяч и миллионов лет, как, например, эволюция человека.
В результате микроэволюции формируется все многообразие видов живых организмов, когда-либо существовавших и ныне живущих на Земле.
Вместе с тем эволюция необратима, и уже исчезнувшие виды никогда не возникают вновь. Появляющиеся виды закрепляют все достигнутое в процессе эволюции, однако это не гарантирует того, что в будущем не появятся новые виды, которые будут иметь более совершенные приспособления к условиям окружающей среды.
Образование новых видов
В широком смысле под образованием новых видов понимается не только отщепление от основного ствола вида нового или распад материнского вида на несколько дочерних, а и общее развитие вида как целостной системы, приводящее к существенным изменениям его морфоструктурной организации. Однако чаще все же видообразование рассматривают как процесс формирования новых видов посредством разветвления «родословного древа» вида. Принципиальное решение проблемы видообразования было предложено Ч. Дарвином. Согласно его теории расселение особей одного вида приводит к образованию популяций, которые вследствие различий условий окружающей среды вынуждены адаптироваться к ним. Это, в свою очередь, влечет за собой обострение внутривидовой борьбы за существование, направляемой естественным отбором. В настоящее время считается, что борьба за существование вовсе не является обязательным фактором видообразования, наоборот, давление отбора в ряде популяций может снижаться. Различие условий существования способствует возникновению неодинаковых приспособительных изменений в популяциях вида, следствием которого является расхождение признаков и свойств популяций —дивергенция. Однако накопления различий, даже на генетическом уровне, отнюдь не достаточно для появления нового вида. До тех пор, пока различающиеся по каким-либо признакам популяции не только контактируют, но и способны к скрещиванию с образованием плодовитого потомства, они относятся к одному виду. Лишь невозможность перетока генов из одной группы особей в другую, даже в случае разрушения разделяющих их преград, т. е. скрещивания, означает завершение сложнейшего эволюционного процесса формирования нового вида.
Видообразование является продолжением микроэволюционных процессов. Существует точка зрения, что видообразование не может быть сведено к микроэволюции, оно представляет качественный этап эволюции и осуществляется благодаря другим механизмам.
Способы видообразования
Выделяют два основных способа видообразования: аллопатрическое и симпатрическое.
Аллопатрическое, или географическое видообразование является следствием пространственного разделения популяций физическими преградами (горные хребты, моря и реки) вследствие их возникновения или расселения в новые места обитания (географическая изоляция). Поскольку в данном случае генофонд отделившейся популяции существенно отличается от материнской, да и условия в месте ее обитания не будут совпадать с исходными, со временем это приведет к дивергенции и формированию нового вида. Ярким примером географического видообразования является обнаруженное Ч. Дарвином во время путешествия на корабле «Бигль» разнообразие видов вьюрков на Галапагосских островах вблизи побережья Эквадора. По-видимому, отдельные особи единственного обитающего на южноамериканском континенте вьюрка каким-то образом попали на острова, и, вследствие различия условий (в первую очередь доступности пищи) и географической изоляции, они постепенно эволюционировали, образовав группу родственных видов. В основе симпатрического, или биологического видообразования лежит какая-либо из форм репродуктивной изоляции, при этом новые виды возникают внутри ареала исходного вида. Обязательным условием симпатрического видообразования является быстрая изоляция образовавшихся форм. Это более быстрый процесс, чем аллопатрическое видообразование, а новые формы похожи на исходные предковые.
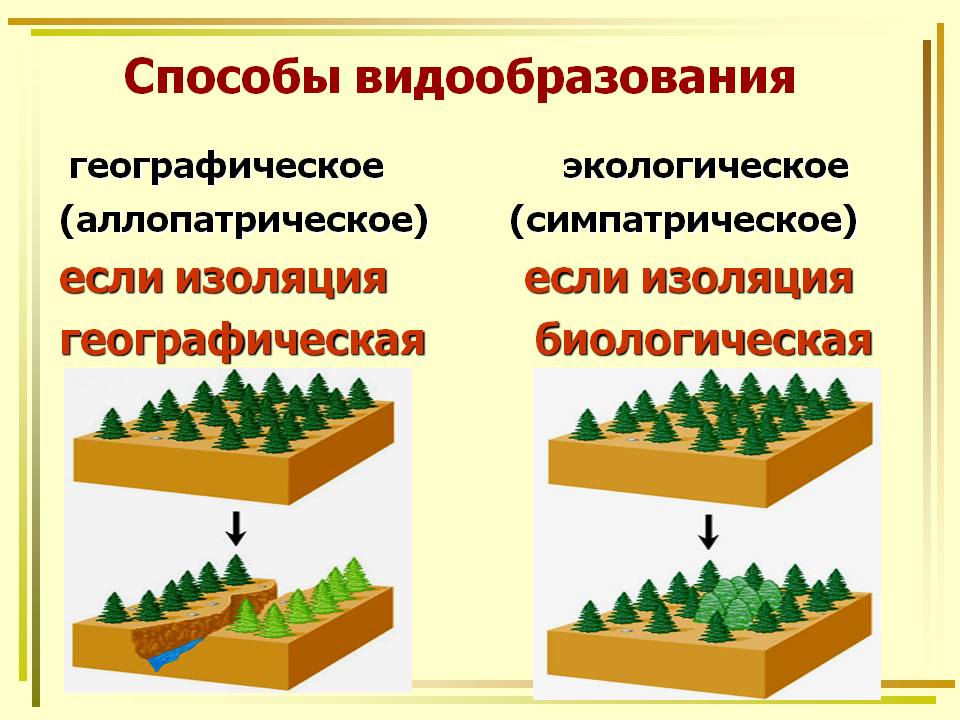
Симпатрическое видообразование может быть вызвано быстрыми изменениями хромосомного набора (полиплоидизация) или хромосомными перестройками. Иногда новые виды возникают вследствие гибридизации двух исходных видов, как, например, у домашней сливы, являющейся гибридом терна и алычи. В некоторых случаях симпатрическое видообразование связано с раз-делением экологических ниш у популяций одного вида в пределах единого ареала или сезонной изоляции — расхождения сроков репродукции у растений (разные виды сосны в Калифорнии пылят в феврале и апреле) и сроков размножения у животных.
Из всего многообразия вновь возникающих видов только немногие, наиболее приспособленные, могут существовать длительное время и дать начало новым видам. Причины гибели большинства видов до сих пор неизвестны, скорее всего это обусловлено резкими изменениями климата, геологическими процессами и вытеснением их более приспособленными организмами. В настоящее время одной из причин гибели значительного числа видов является человек, который истребляет наиболее крупных животных и самые красивые растения, и если в XVII веке этот процесс только начался истреблением последнего тура, то в XXI веке ежечасно исчезает более 10 видов. |


 Меню сайта
Меню сайта Категории раздела
Категории раздела Вход
Вход Поиск
Поиск Друзья сайта
Друзья сайта Наш опрос
Наш опрос Статистика
Статистика
