Блок 3. Организм как биологическая система
3.5. Закономерности наследственности, их цитологические основы. Закономерности наследования, установленные Г. Менделем, их цитологические основы (моно- и дигибридное скрещивание). Законы Т. Моргана: сцепленное наследование признаков, нарушение сцепления генов. Генетика пола. Наследование признаков, сцепленных с полом. Взаимодействие генов. Генотип как целостная система. Генетика человека. Методы изучения генетики человека. Решение генетических задач. Составление схем скрещивания.
Закономерности наследственности, их цитологические основы
Согласно хромосомной теории наследственности каждая пара генов локализована в паре гомо-логичных хромосом, причем каждая из хромосом несет только по одному из этих факторов. Если представить, что гены являются точечными объектами на прямых — хромосомах, то схематически гомозиготные особи могут быть записаны как А||А или а||а, тогда как гетерозиготная — А||а. При образовании гамет в процессе мейоза каждый из генов пары гетерозиготы окажется в одной из половых клеток.
Например, если скрестить двух гетерозиготных особей, то при условии образования у каждой из них только пары гамет возможно получение всего лишь четырех дочерних организмов, три из которых будут нести хотя бы один доминантный ген А, и только один будет гомозиготен по рецессивному гену а, т. е. закономерности наследственности носят статистический характер.
В тех случаях, если гены располагаются в разных хромосомах, то при образовании гамет распределение между ними аллелей из данной пары гомологичных хромосом происходит совершенно независимо от распределения аллелей из других пар. Именно случайное расположение гомологичных хромосом на экваторе веретена в метафазе I мейоза и их последующее расхождение в анафазе I ведет к разнообразию рекомбинаций аллелей в гаметах.
Число возможных сочетаний аллелей в мужских или женских гаметах можно определить по общей формуле 2n, где n — число хромосом, характерное для гаплоидного набора. У человека n = 23, а возможное число сочетаний составляет 223 = 8388608. Последующее объединение гамет при оплодотворении является также случайным, и поэтому в потомстве можно зафиксировать независимое расщепление по каждой паре признаков.
Однако число признаков у каждого организма во много раз больше числа его хромосом, которые можно различить под микроскопом, следовательно, каждая хромосома должна содержать множество факторов. Если представить себе, что у некоторой особи, гетерозиготной по двум парам генов, расположенных в гомологичных хромосомах, образуются гаметы, то следует учитывать не только вероятность образования гамет с исходными хромосомами, но и гамет, получивших измененные в результате кроссинговера в профазе I мейоза хромосомы. Следовательно, в потомстве возникнут новые сочетания признаков. Данные, полученные в экспериментах на дрозофиле, легли в основу хромосомной теории наследственности.
Другое фундаментальное подтверждение цитологической основы наследственности было получено при исследовании различных заболеваний. Так, у человека одна из форм рака обусловлена утратой маленького участка одной из хромосом.
Закономерности наследования, установленные Г. Менделем, их цитологические основы (моно- и дигибридное скрещивание)
Основные закономерности независимого наследования признаков были открыты Г. Менделем, который достиг успеха, применив в своих исследованиях новый на тот момент гибридологический метод.
Успех Г. Менделя был обеспечен следующими факторами:
1. удачным выбором объекта исследования (гороха посевного), который имеет короткий срок вегетации, является самоопыляемым растением, дает значительное количество семян и представлен большим количеством сортов с хорошо различимыми признаками;
2. использованием только чистых линий гороха, которые на протяжении нескольких поколений не давали расщепления признаков в потомстве;
3. концентрацией только на одном-двух признаках;
4. планированием эксперимента и составлением четких схем скрещивания;
5. точным количественным подсчетом полученного потомства.
Для исследования Г. Мендель отобрал только семь признаков, имеющих альтернативные (контрастные) проявления. Уже в первых скрещиваниях он обратил внимание, что в потомстве первого поколения при скрещивании растений с желтыми и зелеными семенами все потомство имело желтые семена. Аналогичные результаты были получены и при исследовании других признаков. Признаки, которые преобладали в первом поколении, Г. Мендель назвал доминантными. Те же из них, которые не проявлялись в первом поколении, получили название рецессивных.
Особи, которые давали расщепление в потомстве, получили название гетерозиготных, а особи, не дававшие расщепления, —гомозиготных.
Признаки гороха, наследование которых изучено Г. Менделем
| Признак |
Вариант проявления |
| Доминантный |
Рецессивный |
| Окраска семян |
Желтая |
Зеленая |
| Форма семян |
Гладкая |
Морщинистая |
| Форма плода (боба) |
Простой |
Членистый |
| Окраска плода |
Зеленая |
Желтая |
| Окраска венчика цветка |
Красная |
Белая |
| Положение цветков |
Пазушные |
Верхушечные |
| Длина стебля |
Длинный |
Короткий |
Скрещивание, при котором исследуется проявление только одного признака, называется моно-гибридным. В таком случае прослеживаются закономерности наследования только двух вариантов одного признака, развитие которых обусловлено парой аллельных генов. Например, признак «окраска венчика цветка» у гороха имеет только два проявления — красная и белая. Все остальные признаки, свойственные данным организмам, во внимание не принимаются и не учитываются в расчетах.
Схема моногибридного скрещивания такова:
 Скрестив два растения гороха, одно из которых имело желтые семена, а другое — зеленые, в первом поколении Г. Мендель получал растения исключительно с желтыми семенами, независимо от того, какое растение было выбрано в качестве материнского, а какое — отцовского. Такие же результаты были получены и в скрещиваниях по другим признакам, что дало Г. Менделю основания сформулировать закон единообразия гибридов первого поколения, который также называют первым законом Менделя и законом доминирования.
Первый закон Менделя: При скрещивании гомозиготных родительских форм, отличающихся по одной паре альтернативных признаков, все гибриды первого поколения будут единообразны как по генотипу, так и по фенотипу.
А — желтые семена; а — зеленые семена.
При самоопылении (скрещивании) гибридов первого поколения оказалось, что 6022 семени имеют желтую окраску, а 2001 — зеленую, что примерно соответствует соотношению 3:1. Обнаруженная закономерность получила название закона расщепления, или второго закона Менделя.
Второй закон Менделя: При скрещивании гетерозиготных гибридов первого поколения в потомстве будет наблюдаться преобладание одного из признаков в соотношении 3:1 по фенотипу (1:2:1 по генотипу).
Однако по фенотипу особи далеко не всегда удается установить ее генотип, поскольку как гомозиготы по доминантному гену (АА), так и гетерозиготы (Аа) будут иметь в фенотипе проявление доминантного гена. Поэтому для организмов с перекрестным оплодотворением применяют анализирующее скрещивание — скрещивание, при котором организм с неизвестным генотипом скрещивается с гомозиготой по рецессивному гену для проверки генотипа. При этом гомозиготные особи по доминантному гену расщепления в потомстве не дают, тогда как в потомстве гетерозиготных наблюдается равное количество особей как с доминантным, так и с рецессивным признаками:
Основываясь на результатах собственных экспериментов, Г. Мендель предположил, что наследственные факторы при образовании гибридов не смешиваются, а сохраняются в неизменном виде. Поскольку связь между поколениями осуществляется через гаметы, то он допустил, что в процессе их образования в каждую из гамет попадает только один фактор из пары (т. е. гаметы генетически чисты), а при оплодотворении пара восстанавливается. Эти предположения получили название правила чистоты гамет.
Правило чистоты гамет: При гаметогенезе гены одной пары разделяются, т. е. каждая гамета несет только один вариант гена.
Однако организмы отличаются друг от друга по многим признакам, поэтому установить закономерности их наследования возможно только при анализе двух и более признаков в потомстве. Скрещивание, при котором рассматривается наследование и производится точный количественный учет потомства по двум парам признаков, называется дигибридным. Если же анализируется проявление большего числа наследственных признаков, то это уже полигибридное скрещивание.
Схема дигибридного скрещивания:
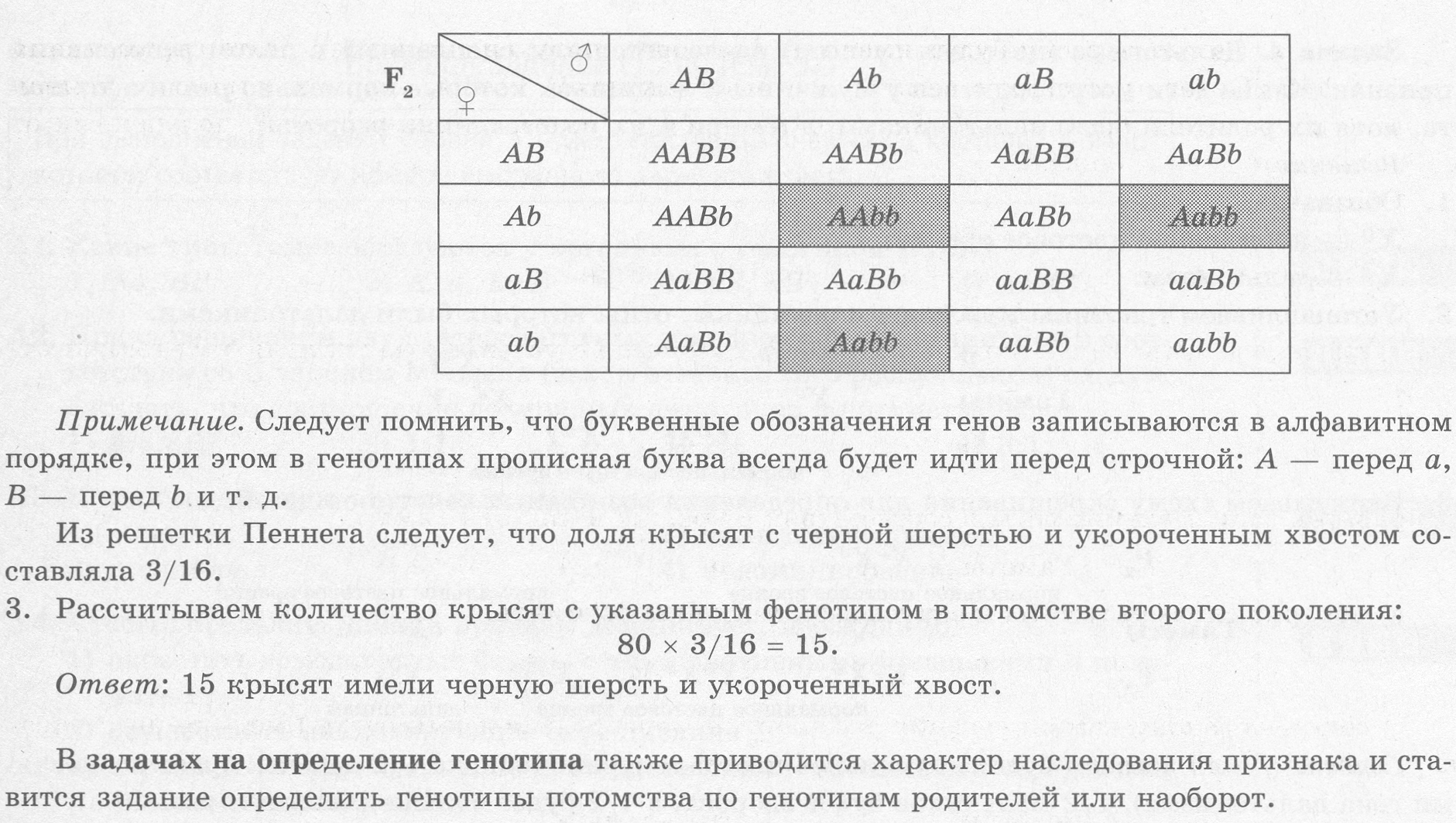
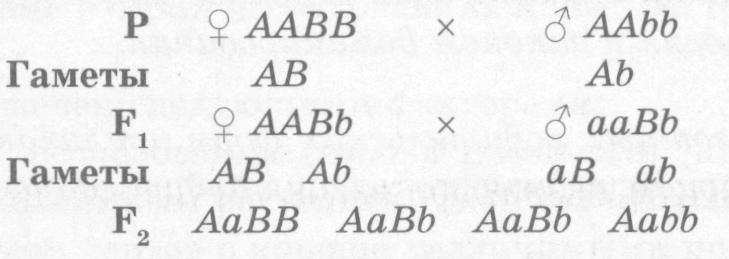 При большем разнообразии гамет определение генотипов потомков становится затруднительным, поэтому для анализа широко используется решетка Пеннета, в которую по горизонтали заносятся мужские гаметы, а по вертикали - женские. Генотипы потомков определяются сочетанием генов в столбцах и строках.
Третий закон Менделя: При скрещивании гомозиготных родительских форм, отличающихся по двум и более парам признаков, во втором поколении будет происходить независимое расщепление данных признаков в соотношении 3:1 (9:3:3:1 при дигибридном скрещивании).
Третий закон Менделя применим только к случаям независимого наследования, когда гены расположены в разных парах гомологичных хромосом. В тех случаях, когда гены расположены в одной паре гомологичных хромосом, действительны закономерности сцепленного наследования. Закономерности независимого наследования признаков, установленные Г. Менделем, также часто нарушаются и при взаимодействии генов.
Законы Т. Моргана: сцепленное наследование признаков, нарушение сцепления генов
Новый организм получает от родителей не россыпь генов, а целые хромосомы, при этом количество признаков и соответственно определяющих их генов гораздо больше, чем хромосом. В соответствии с хромосомной теорией наследственности, гены, расположенные в одной хромосоме, наследуются сцепленно. Вследствие этого при дигибридном скрещивании они не дают ожидаемого расщепления 9:3:3:1 и не подчиняются третьему закону Менделя. Можно было бы ожидать, что сцепление генов является полным, и при скрещивании гомозиготных по данным генам особей и во втором поколении дает исходные фенотипы в соотношении 3:1, а при анализирующем скрещивании гибридов первого поколения расщепление должно составлять 1:1.
Для проверки этого предположения американский генетик Т. Морган выбрал у дрозофилы пару генов, контролирующих окраску тела (серое — черное) и форму крыла (длинные — зачаточные), которые расположены в одной паре гомологичных хромосом. Серое тело и длинные крылья являются доминантными признаками. При скрещивании гомозиготной мухи с серым телом и длинными крыльями и гомозиготной мухи с черным телом и зачаточными крыльями во втором поколении действительно были получены в основном родительские фенотипы в соотношении, близком к 3:1, однако имелось и незначительное количество особей с новыми комбинациями этих признаков.
Данные особи называются рекомбинантными. Однако, проведя анализирующее скрещивание гибридов первого поколения с гомозиготами по рецессивным генам, Т. Морган обнаружил, что 41,5% особей имели серое тело и длинные крылья, 41,5% — черное тело и зачаточные крылья, 8,5% — серое тело и зачаточные крылья, и 8,5% — черное тело и зачаточные крылья. Он связал полученное расщепление с кроссинговером, происходящим в профазе I мейоза и предложил считать единицей расстояния между генами в хромосоме 1 % кроссинговера, впоследствии названный в его честь морганидой.
Закономерности сцепленного наследования, установленные в ходе экспериментов на дрозофиле, получили название закона Т. Моргана.
Закон Моргана: Гены, локализованные в одной хромосоме, занимают определенное место, называемое локусом, и наследуются сцепленно, причем сила сцепления обратно пропорциональна расстоянию между генами.
Гены, расположенные в хромосоме непосредственно друг за другом (вероятность кроссинговера крайне мала), называются сцепленными полностью, а если между ними находится еще хотя бы один ген, то они сцеплены не полностью и их сцепление нарушается при кроссинговере в резуль¬тате обмена участками гомологичных хромосом.
Явления сцепления генов и кроссинговера позволяют построить карты хромосом с нанесенным на них порядком расположения генов. Генетические карты хромосом созданы для многих генетически хорошо изученных объектов: дрозофилы, мыши, человека, кукурузы, пшеницы, гороха и др. Изучение генетических карт позволяет сравнивать строение генома у различных видов организмов, что имеет важное значение для генетики и селекции, а также эволюционных исследований.
Генетика пола
Пол — это совокупность морфологических и физиологических особенностей организма, обе-спечивающих половое размножение, сущность которого сводится к оплодотворению, то есть слиянию мужских и женских половых клеток в зиготу, из которой развивается новый организм.
Признаки, по которым один пол отличается от другого, делят на первичные и вторичные. К первичным половым признакам относятся половые органы, а все остальные — ко вторичным.
У человека вторичными половыми признаками являются тип телосложения, тембр голоса, преобладание мышечной или жировой ткани, наличие оволосения на лице, кадыка, молочных желез. Так, у женщин таз обычно шире плеч, преобладает жировая ткань, выражены молочные железы, голос — высокий. Мужчины же отличаются от них более широкими плечами, преобладанием мышечной ткани, наличием оволосения на лице и кадыка, а также низким голосом. Человечество издавна интересовал вопрос, почему особи мужского и женского пола рождаются в соотношении приблизительно 1:1. Объяснение этому было получено при изучении кариотипов насекомых. Оказалось, что у самок некоторых клопов, кузнечиков и бабочек на одну хромосому больше, чем у самцов. В свою очередь самцы продуцируют гаметы, различающиеся по количеству хромосом, тем самым заранее определяя пол потомка. Однако впоследствии было выяснено, что у большинства организмов количество хромосом у самцов и самок все же не различается, но у одного из полов есть пара хромосом, которые не подходят друг другу по размерам, а у другого все хромосомы парные.
В кариотипе человека также обнаружили подобное различие: у мужчин есть две непарные хромосомы. По форме эти хромосомы в начале деления напоминают латинские буквы X и Y, и поэтому были названы Х- и Y-хромосомами. Сперматозоиды мужчины могут нести одну из этих хромосом и определять пол будущего ребенка. В связи с этим хромосомы человека и многих других организмов делят на две группы: аутосомы и гетерохромосомы, или половые хромосомы.
К аутосомам относят хромосомы, одинаковые для обоих полов, тогда как половые хромосомы — это хромосомы, отличающиеся у разных полов и несущие информацию о половых признаках. В тех случаях, когда пол несет одинаковые половые хромосомы, например XX, говорят, что он гомозиготен, или гомогаметен (образует одинаковые гаметы). Другой же пол, имеющий разные половые хромосомы (XY), называется гемизиготным (не имеющим полного эквивалента аллельных генов), или гетерогаметным. У человека, большинства млекопитающих, мушки дрозофилы и других организмов гомогаметен женский пол (XX), а мужской — гетерогаметен (XY), тогда как у птиц гомогаметен мужской пол (ZZ, или XX), а женский — гетерогаметен (ZW, или XY).
Х-хромосома является крупной неравноплечей хромосомой, которая несет свыше 1500 генов, причем многие мутантные их аллели вызывают у человека развитие тяжелых наследственных заболеваний, таких как гемофилия и дальтонизм. Y-хромосома, напротив, очень маленькая, в ней содержится всего около десятка генов, в том числе специфические гены, ответственные за развитие по мужскому типу.
Кариотип мужчины записывается как ♂46, XY, а кариотип женщины — как ♀ 46, XX.
Поскольку гаметы с половыми хромосомами продуцируются у самцов с равной вероятностью, то ожидаемое соотношение полов в потомстве составляет 1:1, что и совпадает с фактически наблюдаемым.
Пчелы отличаются от других организмов тем, что самки у них развиваются из оплодотворенных яиц, а самцы — из неоплодотворенных. Соотношение полов у них отличается от указанного выше, так как процесс оплодотворения регулирует матка, в половых путях которой с весны запасаются сперматозоиды на весь год.
У ряда организмов пол может определяться иным способом: до оплодотворения или после него, в зависимости от условий внешней среды.
Наследование признаков, сцепленных с полом
Так как некоторые гены находятся в половых хромосомах, неодинаковых у представителей противоположных полов, то характер наследования признаков, кодируемых данными генами, отличается от общего. Такой тип наследования называется крис-кросс наследованием, поскольку мужчины наследуют признаки матери, а женщины — отца. Признаки, определяемые генами, которые находятся в половых хромосомах, называются сцепленными с полом. Примерами признаков, сцепленных с полом, являются рецессивные признаки гемофилии и дальтонизма, которые в основном проявляются у мужчин, так как в Y-хромосоме нет аллельных генов. Женщины болеют такими болезнями только в том случае, если и от отца, и от матери они получили такие признаки.
Например, если мать была гетерозиготным носителем гемофилии, то у половины ее сыновей свертываемость крови будет нарушена: Хн— нормальное свертывание крови Xh — несвертываемость крови (гемофилия)
Признаки, закодированные в генах Y-хромосомы, передаются сугубо по мужской линии и называются голандрическими (наличие перепонки между пальцами ног, повышенное оволосение края ушной раковины).
Взаимодействие генов
Проверка закономерностей независимого наследования на различных объектах уже в начале XX века показала, что, например, у ночной красавицы при скрещивании растений с красным и белым венчиком у гибридов первого поколения венчики окрашены в розовый цвет, тогда как во втором поколении имеются особи с красными, розовыми и белыми цветками в соотношении 1:2:1. Это навело исследователей на мысль, что аллельные гены могут оказывать определенное влияние друг на друга. Впоследствии было также установлено, что и неаллельные гены способствуют проявлению признаков других генов или подавляют их. Данные наблюдения стали основой представления о генотипе как о системе взаимодействующих генов. В настоящее время различают взаимодействие аллельных и неаллельных генов.
Ко взаимодействию аллельных генов относят полное и неполное доминирование, кодоминирование и сверхдоминирование. Полным доминированием считают все случаи взаимодействия аллельных генов, при которых в гетерозиготе наблюдается проявление исключительно доминантного признака, как, например, окраска и форма семени у гороха.
Неполное доминирование — это тип взаимодействия аллельных генов, при котором проявление рецессивного аллеля в большей или меньшей степени ослабляет проявление доминантного, как в случае окраски венчика ночной красавицы (белая + красная = розовая) и шерсти у крупного рогатого скота.
Кодоминированием называют такой тип взаимодействия аллельных генов, при котором оба аллеля проявляются, не ослабляя эффектов друг друга. Типичным примером кодоминирования является наследование групп крови по системе ABO (табл. 3.2). IV (АВ) группа крови у человека (генотип — IАIВ).
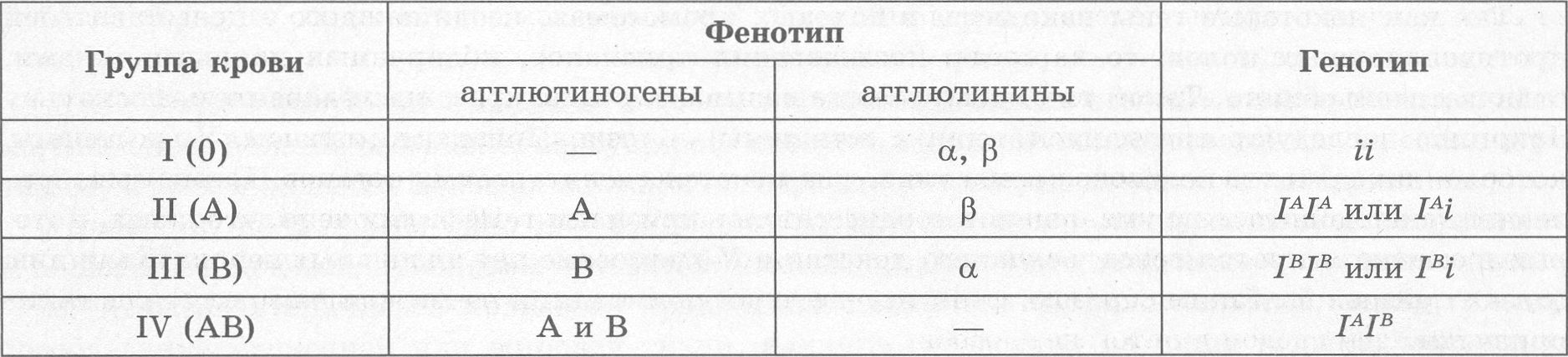
Как видно из таблицы, I, II и III группы крови наследуются по типу полного доминирования, тогда как IV (АВ) группа (генотип — IАIВ) является случаем кодоминирования.
Особым случаем взаимодействия аллельных генов можно считать так называемые летальные гены, которые в гомозиготном состоянии приводят к гибели организма чаще всего в эмбриональном периоде. Причиной гибели потомства является плейотропное действие генов серой окраски шерсти у каракулевых овец, платиновой окраски у лис и отсутствие чешуи у зеркальных карпов. При скрещивании двух гетерозиготных по этим генам особей расщепление по исследуемому признаку в потомстве будет равняться 2:1 вследствие гибели 1/4 потомства.
Основными типами взаимодействия неаллельных генов являются комплементарность, эпистаз и полимерия. Комплементарность — это тип взаимодействия неаллельных генов, при котором для проявления определенного состояния признака необходимо присутствие как минимум двух доминантных аллелей разных пар. Например, у тыквы при скрещивании растений со сферически-ми (ААbb) и длинными (ааВВ) плодами в первом поколении появляются растения с дисковидными плодами (АаВb).
К эпистазу относят такие явления взаимодействия неаллельных генов, при которых один неаллельный ген подавляет развитие признака другого. Например, у кур окраска оперения определяется одним доминантным геном, тогда как другой доминантный ген подавляет развитие окраски, в результате чего большинство кур имеет белое оперение.
Полимерией называют явление, при котором неаллельные гены оказывают одинаковое влияние на развитие признака. Таким образом чаще всего кодируются количественные признаки. Например, цвет кожи человека определяется как минимум четырьмя парами неаллельных генов — чем больше доминантных аллелей в генотипе, тем темнее кожа.
Генотип как целостная система
Генотип не является механической суммой генов, поскольку возможность проявления гена и форма его проявления зависят от условий среды. В данном случае под средой понимается не только окружающая среда, но и генотипическая среда — другие гены.
Проявление качественных признаков редко зависит от условий окружающей среды, хотя, если у горностаевого кролика выбрить участок тела с белой шерстью и прикладывать к нему пузырь со льдом, то со временем на этом месте вырастет черная шерсть.
Развитие количественных признаков намного сильнее зависит от условий окружающей среды. Например, если современные сорта пшеницы возделывать без применения минеральных удобрений, то ее урожайность будет существенно отличаться от генетически запрограммированных 100 и более центнеров с гектара.
Таким образом, в генотипе записаны лишь «способности» организма, однако проявляются они только во взаимодействии с условиями окружающей среды.
Кроме того, гены взаимодействуют друг с другом и, оказавшись в одном генотипе, могут сильно влиять на проявление действия соседних генов. Таким образом, для каждого отдельно взятого гена существует генотипическая среда. Возможно, что развитие любого признака связано с действием многих генов. Кроме того, выявлена зависимость нескольких признаков от одного гена. Например, у овса окраска чешуек и длина ости семени определяются одним геном. У дрозофилы ген белой окраски глаза одновременно влияет на цвет тела и внутренних органов, длину крыльев, снижение плодовитости и уменьшение продолжительности жизни. Не исключено, что каждый ген одновременно является геном основного действия для «своего» признака и модификатором для других признаков. Таким образом, фенотип — это результат взаимодействия генов всего генотипа с окружающей средой в онтогенезе особи.
В связи с этим известный российский генетик М. Е. Лобашев определил генотип как систему взаимодействующих генов. Сложилась эта целостная система в процессе эволюции органического мира, при этом выживали лишь те организмы, у которых взаимодействие генов давало наиболее благоприятную реакцию в онтогенезе.
Генетика человека
Для человека как биологического вида в полной мере справедливы генетические закономерности наследственности и изменчивости, установленные для растений и животных. Вместе с тем генетика человека, изучающая закономерности наследственности и изменчивости у человека на всех уровнях его организации и существования, занимает особое место среди других разделов генетики.
Генетика человека одновременно является фундаментальной и прикладной наукой, поскольку занимается исследованием наследственных болезней человека, которых в настоящее время описано уже более 4 тыс. Она стимулирует развитие современных направлений общей и молекулярной генетики, молекулярной биологии и клинической медицины. В зависимости от проблематики генетика человека делится на несколько направлений, резвившихся в самостоятельные науки: генетика нормальных признаков человека, медицинская генетика, генетика поведения и интеллекта, популяционная генетика человека. В связи с этим в наше время человек как генетический объект исследован едва ли не лучше, чем основные модельные объекты генетики: дрозофила, арабидопсис и др.
Биосоциальная природа человека накладывает значительный отпечаток на исследования в области его генетики вследствие позднего полового созревания и больших временных разрывов между поколениями, малочисленности потомства, невозможности направленных скрещиваний для генетического анализа, отсутствия чистых линий, недостаточной точности регистрации наследственных признаков и небольших родословных, невозможности создания одинаковых и строго контролируемых условий для развития потомков от разных браков, сравнительно большого числа плохо различающихся хромосом и невозможности экспериментального получения мутаций.
Методы изучения генетики человека
Методы, применяемые в генетике человека, принципиально не отличаются от общепринятых для других объектов — это генеалогический, близнецовый, цитогенетический, дерматоглифический, молекулярно-биологический и популяционно-статистический методы, метод гибридизации соматических клеток и метод моделирования. Их использование в генетике человека учитывает специфику человека как генетического объекта.
Близнецовый метод помогает определить вклад наследственности и влияние условий окружающей среды на проявление признака на основе анализа совпадения этих признаков у однояйцевых и разнояйцевых близнецов. Так, у большинства однояйцевых близнецов совпадают группы крови, цвет глаз и волос, а также целый ряд других признаков, тогда как корью болеют одновременно оба типа близнецов.
Дерматоглифический метод основан на исследовании индивидуальных особенностей кожных рисунков пальцев рук (дактилоскопия), ладоней и ступней ног. На основе этих особенностей он зачастую позволяет своевременно выявить наследственные заболевания, в частности хромосомные аномалии, такие как синдром Дауна, Шерешевского - Тернера и др.
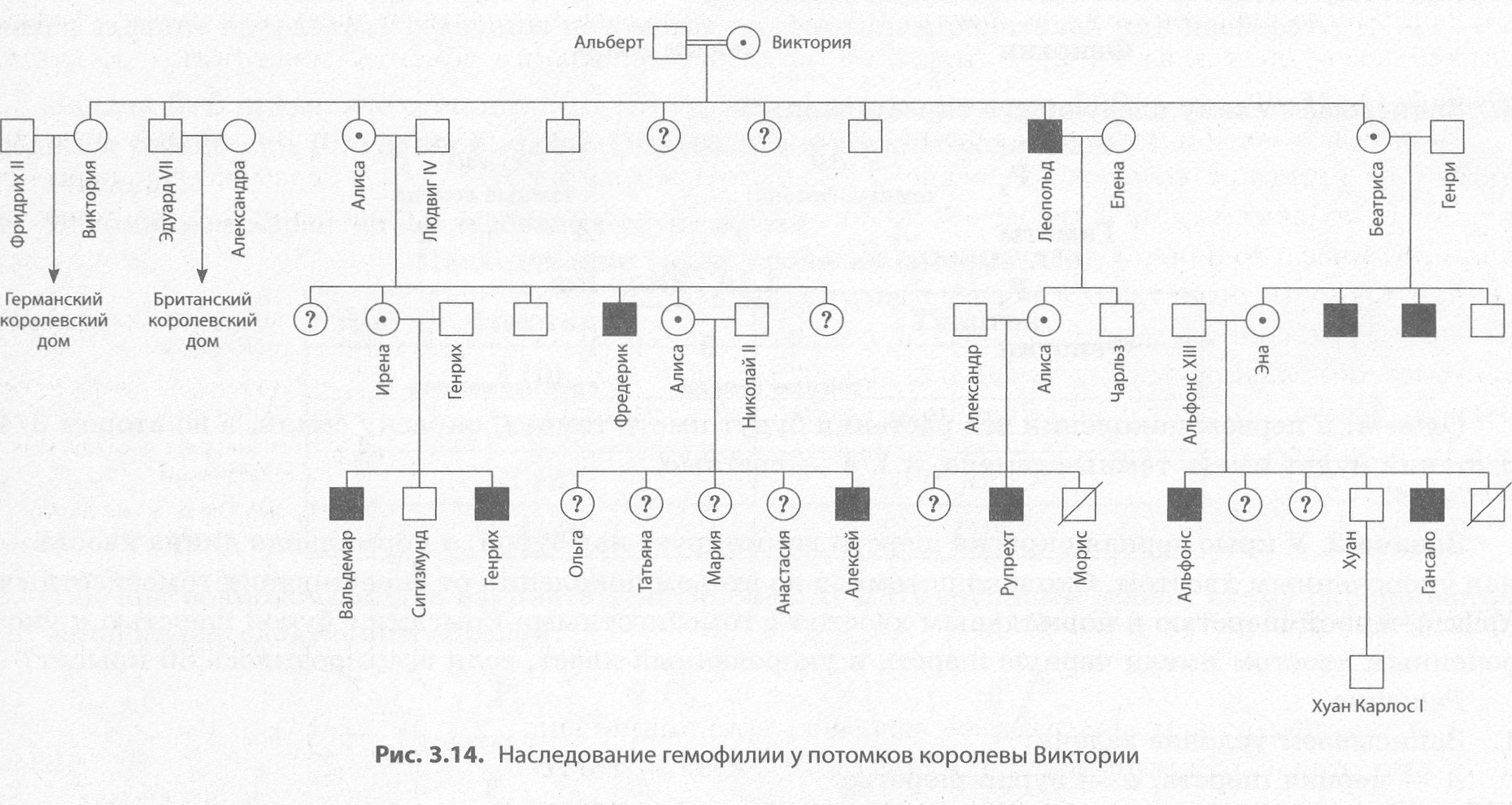
Генеалогический метод — это метод составления родословных, с помощью которых определяют характер наследования изучаемых признаков, в том числе наследственных болезней, и прогнозируют рождение потомков с соответствующими признаками. Он позволил выявить наследственную природу таких заболеваний, как гемофилия, дальтонизм, хорея Гентингтона и др. еще до открытия основных закономерностей наследственности. При составлении родословных ведут записи о каждом из членов семьи и учитывают степень родства между ними. Далее на основании полученных данных с помощью специальной символики строится родословное древо.
Генеалогический метод можно использовать на одной семье, если есть сведения о достаточном количестве прямых родственников человека, родословная которого составляется — пробанда, — по отцовской и материнской линиям, в противном случае собирают сведения о нескольких семьях, в которых проявляется данный признак. Генеалогический метод позволяет установить не только наследуемость признака, но и характер наследования: доминантный или рецессивный, аутосомный или сцепленный с полом и т. д. Так, по портретам австрийских монархов Габсбургов было установлено наследование прогнатии (сильно выпяченной нижней губы) и «королевской гемофилии» у потомков британской королевы Виктории.
 Решение генетических задач. Составление схем скрещивания
Решение генетических задач. Составление схем скрещивания
Все разнообразие генетических задач можно свести к трем типам:
1. Расчетные задачи.
2. Задачи на определение генотипа.
3. Задачи на установление типа наследования признака.
Особенностью расчетных задач является наличие информации о наследовании признака и фенотипах родителей, по которым легко установить и генотипы родителей. В них требуется установить генотипы и фенотипы потомства.
|


 Меню сайта
Меню сайта Категории раздела
Категории раздела Вход
Вход Поиск
Поиск Друзья сайта
Друзья сайта Наш опрос
Наш опрос Статистика
Статистика